Archives
- 2026-07
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
The heterogeneity among studies that investigated the associ
2020-10-10
The heterogeneity among studies that investigated the association of COMT Val158Met with alexithymia was also significant (Ham et al., 2005; Min et al., 2016; Swart et al., 2011; Zekioglu et al., 2014). The healthy status, drug effect, and other demographic variables also influenced one\'s ability o
-
liothyronine sodium Protein kinase casein kinase CK comprise
2020-10-10
Protein kinase casein kinase 1 (CK1) comprises a family of highly related, constitutively active serine/threonine protein kinases (reviewed by [15]). CK1 is involved in controlling a wide variety of different cellular events including protein turnover [16], [17], nuclear import [18] and the cellular
-
In ER group Histopathological examination followed by Fluore
2020-10-09
In ER + group, Histopathological examination followed by Fluorescence in situ hybridization (FISH) had revealed the absence of HER2 receptors. Co-targeting of ER and HER2 appears to provide benefit without a significant increase in toxicity although formal trials have not been carried out [18]. Adop
-
The role of Epac in the regulation of intracellular Ca
2020-10-09
The role of Epac in the regulation of intracellular Ca homeostasis and contractility is still matter of debate. In rat adult cardiomyocytes, acute Epac stimulation decreased the amplitude of Ca transients [6], [9], [10] with either no changes [6] or increments [9] in cell shortening, suggesting an e
-
An important focus of the present study
2020-10-09
An important focus of the present study has been the interaction of HUVECs with extracellular matrix in the regulation of angiogenesis. Matrigel is a gelatinous protein extract secreted by Engelbreth–Holm–Swarm mouse sarcoma cells, and is considered to be a good model of the extracellular matrix (Kl
-
First of all it should be
2020-10-09
First of all, it should be emphasized that, although cPLA2α manifests a marked selectivity for phospholipids that contain AA at the sn-2 position, this does not mean in any way that, in cells, the enzyme cannot hydrolyze other fatty acyl residues to a significant extent even if this occurs at a lowe
-
Regardless of its effect on
2020-10-09
Regardless of its effect on EBV infection, the chemokine system has been found to be regulated in both nasopharyngeal cancer and Hodgkin lymphoma. As such, Hodgkin and Reed–Sternberg (HRS) cells from Hodgkin lymphoma and nasopharyngeal carcinoma cells have been shown to express CXCL8 and CCL17. Furt
-
Recently a method for in vivo photoactivation of
2020-10-09
Recently, a method for in vivo photoactivation of PEG Virus Precipitation Kit sale expressing PA-GFP in precise microanatomical compartments was described (Victora et al., 2010), which makes it possible to optically mark Tfh cells and track them 20 hr later (Shulman et al., 2013). Unexpectedly, it w
-
Leonurine hydrochloride The majority of APC C subunits form
2020-10-09
The majority of APC/C subunits form a giant scaffold, which was originally named the arc lamp, based on its shape when viewing APC/C from one side (Figure 1 and Box 1) [24]. The scaffold consists of two modules: the TPR lobe resembles the curved post and lamp, and the platform resembles the Leonurin
-
MuRF and MuRF in http www
2020-10-09
MuRF1 and MuRF3 in cooperation with the E2 ubiquitin-conjugating enzymes UbcH5a, -b, and -c were found to mediate degradation of myosin heavy chain β/slow (MHC β/slow) and MHC IIa via UPS, both, in vitro and in vivo [60]. Mice lacking both MuRF1 and MuRF3 developed skeletal muscle myopathy and hyper
-
br Conclusion In the last year
2020-10-09
Conclusion In the last year, significant advances in our understanding of ubiquitin transfer by IAPs and related E3 ligases have been made. The three crystal structures of the RING-bound E2~Ub show that the conjugate is bound by the RING domain in the closed conformation, where the I44-centered f
-
br Regulation of p via the ubiquitin
2020-10-09
Regulation of p53 via the ubiquitin-proteasome pathway Studies into regulation of p53 via PTM processes that involve phosphorylation, acetylation, ubiquitination, SUMOylation, neddylation, and methylation are being increasingly reported [23], [24]. Phosphorylation and acetylation of p53 stimulate
-
br Materials and methods br Results
2020-10-09
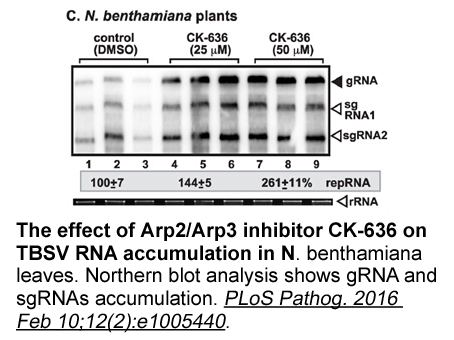
Materials and methods Results Discussion Regardless of obvious beneficial effects of GC on acceleration of foetal lung and cardiomyocytes maturation (Kamath-Rayne et al., 2012; Rog-Zielinska et al., 2015), the current study revealed that GC exposure during late pregnancy induces IR and alte
-
More recently Boskovic et al
2020-10-09
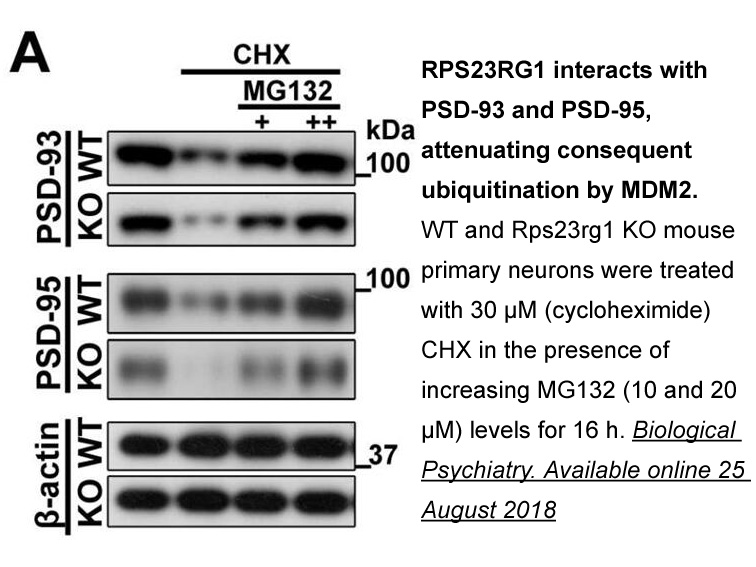
More recently, Boskovic et al. (2003) have visually corroborated that purified DNA-PKcs can bind short DNA fragments by using single particle EM. DNA biotinylated at the 5′ end was labelled by coupling to commercial streptavidin–gold particles (Fig. 4D). When analysed in the microscope, DNA-PKcs mol
-
br Acknowledgements br Introductio http www apexbt
2020-10-09
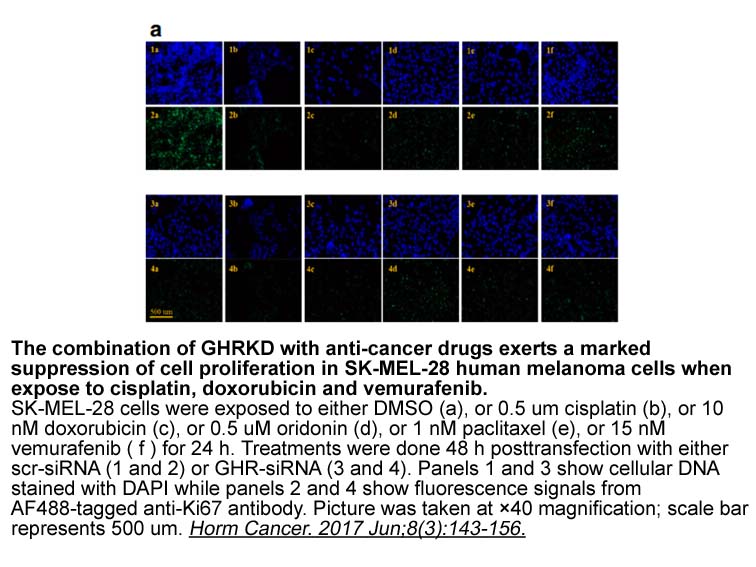
Acknowledgements Introduction TB is also widely prescribed as a stain to identify proteoglycans and glycosaminoglycans (GAGs) in agarose gel electrophoresis (Dietrich and Dietrich, 1976; Coulson-Thomas et al., 2015). In addition, as adjunct to a photodynamic antimicrobial chemotherapy assay, T
16200 records 782/1080 page Previous Next First page 上5页 781782783784785 下5页 Last page